
Stickstoffmanagement 2.0
Stickstoffmanagement 2.0 in der Meeresaquaristik
Meerwasseraquarien sind hochkomplexe, künstliche Ökosysteme, die versuchen, die Nährstoffkreisläufe tropischer Korallenriffe nachzubilden. Doch während natürliche Riffe in extrem nährstoffarmen Gewässern gedeihen, kämpfen viele Aquarianer mit instabilen Nährstoffwerten, Algenproblemen, Cyanobakterien oder stagnierendem Korallenwachstum. Die Ursache liegt häufig in einem fundamental falschen Verständnis von Stickstoff.
Korallen sind keine Einzelorganismen, sondern Holobionten – ein funktionelles Netzwerk aus Koralle, Zooxanthellen (Symbiodiniaceae), Bakterien, Archaeen, Pilzen und Viren. Innerhalb dieses Netzwerks spielt Stickstoff eine zentrale Rolle. Er ist nicht nur ein Nährstoff, sondern ein Regulator, der Wachstum, Farbe, Photosyntheseleistung, Stressresistenz, Kalzifikation und Mikrobiom-Stabilität steuert.
Die moderne Forschung zeigt:
Nicht die Menge des Stickstoffs entscheidet über Korallengesundheit, sondern seine Form, sein Fluss und seine interne Verteilung.
Für die Meeresaquaristik bedeutet das einen Paradigmenwechsel.
1. Stickstoff im Korallenholobionten – was Aquarianer wirklich wissen müssen
Stickstoff ist für Korallen ein essenzielles Element, das in nahezu allen biochemischen Prozessen eine Rolle spielt. Doch anders als in klassischen Aquarienmodellen ist Stickstoff im Korallenholobionten kein frei verfügbarer Pool, sondern ein eng kontrollierter, dynamischer Stoffstrom.
1.1 Stickstoff im Korallengewebe
Das Korallengewebe benötigt Stickstoff für:
- die Synthese von Aminosäuren, Proteinen und Enzymen
- die Bildung von Strukturproteinen für Polypen und Skelettmatrix
- Reparaturprozesse und Geweberegeneration
- die Produktion immunrelevanter Moleküle
- die Aufrechterhaltung der extrazellulären Matrix
Ein Stickstoffmangel führt zu:
- Wachstumsstopp
- Verlust von Gewebevolumen
- reduzierter Immunfunktion
- erhöhter Krankheitsanfälligkeit
- Verlust von Farbintensität
Korallen reagieren auf Stickstoffmangel oft subtil:
Polypen bleiben kleiner, Gewebe wirkt „dünn“, Wachstum stagniert.
Viele Aquarianer interpretieren dies fälschlich als „zu wenig Licht“ oder „zu wenig Spurenelemente“.
1.2 Stickstoff in Symbiodiniaceae
Zooxanthellen benötigen Stickstoff für:
- Chlorophyll‑Synthese
- Aufbau photosynthetischer Enzyme
- Zellteilung
- Stickstoffspeicherung in Form organischer Verbindungen
Wichtig:
Die Koralle hält ihre Symbionten absichtlich stickstofflimitiert, um deren Wachstum zu kontrollieren.
Wird diese Limitierung aufgehoben, beginnen die Symbionten, sich schneller zu teilen als die Koralle — ein zentraler Mechanismus vieler Bleaching‑Ereignisse.
1.3 Stickstoff im Mikrobiom
Das Mikrobiom übernimmt:
- die Umsetzung verschiedener Stickstoffformen
- die Stabilisierung der chemischen Mikrozone um den Polyp
- die Produktion stickstoffhaltiger Metabolite
- die Unterstützung der Immunität
- die Kontrolle des pH‑Werts im Gewebe
Das Mikrobiom ist damit ein aktiver Teil des Stickstoffmanagements.
2. DIN vs. DON – warum Aquarien oft gegen die Natur arbeiten

2.1 Riffe sind DON‑dominiert
In natürlichen Riffen stammt der Großteil des Stickstoffs aus:
- Urea
- Aminosäuren
- Peptiden
- gelösten organischen Stickstoffverbindungen
Diese Formen sind:
- biologisch reguliert
- stabil
- effizient verwertbar
- symbiosefreundlich
2.2 Aquarien sind DIN‑dominiert
Aquarien enthalten überwiegend:
- Nitrat
- Nitrit
- Ammonium
Diese Formen sind:
- leicht messbar
- aber biologisch problematisch
- Ursache vieler Instabilitäten
Riffe sind DON‑dominiert – Aquarien DIN‑dominiert. Das ist der Kern des Problems.
3. Stickstoffformen im Aquarium – vollständige Übersicht
3.1 DIN (anorganisch)
DIN ist direkt messbar, aber biologisch nicht optimal.
- Nitrat (NO₃⁻)
ineffizient, stressfördernd, ROS‑erzeugend - Nitrit (NO₂⁻)
toxisch, instabil - Ammonium (NH₄⁺)
bevorzugt, effizient, symbiosefreundlich
3.2 DON (organisch)
DON ist nicht messbar mit Standardtests, aber physiologisch günstig.
- Urea
- Aminosäuren
- Peptide
DON ist die natürliche Stickstoffform tropischer Riffe.
4. Die 1:100‑Regel – woher sie stammt und warum sie im Meerwasseraquarium nicht gilt
Die 1:100‑Regel (NO₃ : PO₄ ≈ 100 : 1) ist eine Fehlinterpretation des Redfield‑Verhältnisses (C:N:P = 106:16:1).
4.1 Was Redfield wirklich beschreibt
Redfield beschreibt:
- die durchschnittliche Zusammensetzung von marinem Phytoplankton
- und die Zusammensetzung des offenen Ozeans
Es ist ein ökologisches Großsystem‑Mittel, kein Zielwert für Korallen oder Aquarien.
4.2 Warum Nitratüberschuss Phosphatlimitierung verursacht
Ein Nitratüberschuss im Meerwasseraquarium führt fast zwangsläufig zu einer Phosphatlimitierung, selbst wenn der absolute Phosphatwert im Wasser nicht extrem niedrig erscheint. Dieser Zusammenhang ist einer der zentralen Mechanismen, die erklären, warum Korallen unter hohen Nitratwerten schlechter wachsen, blasser werden oder sogar bleichen.
4.2.1 Nitrat beschleunigt das Wachstum der Symbiodiniaceae
Nitrat wirkt wie ein Wachstumssignal.
Sobald Nitrat verfügbar ist, erhöhen die Symbionten:
- ihre Zellteilungsrate
- ihren Chlorophyllgehalt
- ihren Stickstoffbedarf
Jede neue Symbiontenzelle benötigt jedoch Phosphat — und zwar mehr, als die Koralle normalerweise bereitstellt.
4.2.2.Phosphat ist der limitierende Faktor für Zellmembranen
Phosphat wird für die Synthese von:
- Phospholipiden
- ATP
- DNA/RNA
- Photosynthese-Enzymen
benötigt. Wenn Nitrat das Wachstum beschleunigt, steigt der Phosphatbedarf sprunghaft an.
4.2.3 Phosphatlimitierung destabilisiert die Thylakoidmembranen
Fehlt Phosphat, ersetzen Symbionten Phospholipide durch sulfonierte Lipide.
Das führt zu:
- instabilen Thylakoidmembranen
- erhöhter ROS-Produktion
- geringerer Hitzetoleranz
- erhöhter Bleaching-Anfälligkeit
4.2.4.Nitrat verschiebt das N:P-Verhältnis
Ein Anstieg von Nitrat bei konstantem Phosphat verschiebt das Verhältnis dramatisch.
Das System wird phosphatlimitiert, selbst wenn PO₄ messbar bleibt.
4.2.5 Bakterien binden Phosphat
Nitratüberschuss fördert heterotrophe Bakterien, die Phosphat in Biomasse binden.
Das führt zu einer sekundären Phosphatlimitierung.
4.2.6 Korallen benötigen selbst mehr Phosphat
Steigt der Stoffwechsel durch Nitrat, steigt auch der Phosphatbedarf der Koralle.
4.2.7 Ergebnis: versteckte Phosphatlimitierung
Der gemessene PO₄-Wert ist nur die Restmenge, nicht die Verfügbarkeit.
Nitratüberschuss erzeugt Phosphatlimitierung — und damit die Grundlage für Bleaching, Wachstumsstopp und Instabilität.
5. Wie Korallen Stickstoff aufnehmen – Mechanismen und Präferenzen
Korallen und ihre Symbionten bevorzugen Ammonium (NH₄⁺) und Urea.
Diese beiden Formen sind energetisch günstig, stabil und biologisch sinnvoll.
5.1 Ammonium – die bevorzugte Form
Ammonium wird:
- direkt eingebaut
- ohne ROS‑Stress verwertet
- energetisch effizient genutzt
- bevorzugt von Symbiodiniaceae aufgenommen
Ammonium ist die natürliche Stickstoffform in der Mikrozone um den Polyp, die durch bakterielle Aktivität entsteht.
5.2 Urea – die natürliche DON‑Form
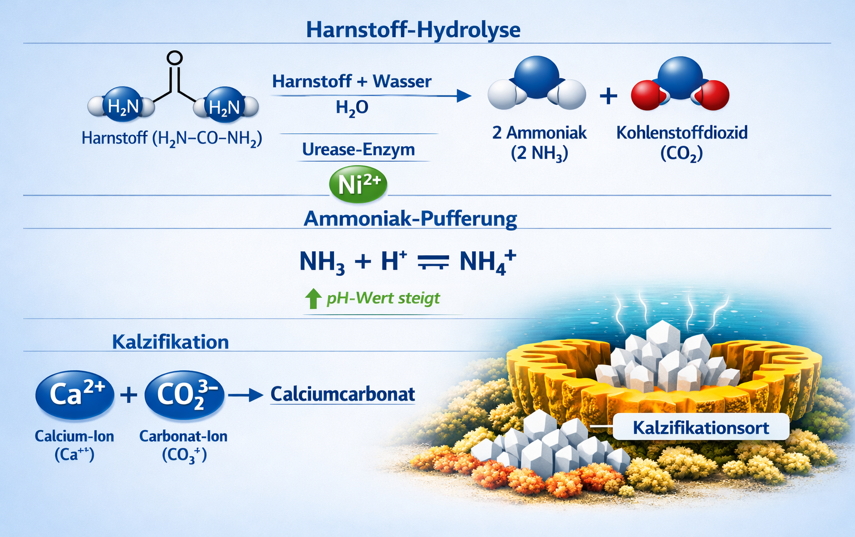
Urea (Harnstoff) wird über Urease zu NH₃ + CO₂ gespalten. Urease ist ein Nickel‑abhängiges Enzym, das in Korallen, Symbiodiniaceae und vielen Riffbakterien vorkommt und die Aufgabe hat, Urea in zwei physiologisch extrem wichtige Moleküle zu spalten: Ammoniak (NH₃) und Kohlendioxid (CO₂).
- NH₃ sofort als Stickstoffquelle für Proteinsynthese, Wachstum und Symbiontenversorgung genutzt wird,
- CO₂ direkt in die Photosynthese einfließt und gleichzeitig die Kalzifikation unterstützt,
- die Reaktion pH‑stabilisierend wirkt, indem NH₃ Protonen bindet,
- Urease selbst nur dann aktiv ist, wenn ausreichend Nickel vorhanden ist.
Urea ist:
- pH‑neutral
- stabil
- stressresistent
- die wichtigste Stickstoffquelle natürlicher Riffe
Urease ist das Schlüsselenzym, das Urea überhaupt erst zu einer wertvollen, stabilen und natürlichen Stickstoffquelle für Korallen macht.
5.3 Nitrat – ineffizient und stressfördernd
Nitrat muss:
- energieaufwendig reduziert werden
- erzeugt ROS
- destabilisiert die Symbiose
6. Stickstoffkreislauf im Aquarium – das Holobiont-Modell
6.1 Der klassische Aquarienkreislauf
Futter → Ammonium → Nitrifikation → Nitrat → Denitrifikation → N₂
6.2 Der natürliche Riffkreislauf
DON → Urea, Aminosäuren, Peptide → direkte Aufnahme → kaum Nitrat
6.3 Ziel im Aquarium
Stickstoff intern recyceln, aber Symbiodinium stickstofflimitiert halten.
Stickstoffmanagement heißt: Fluss steuern, nicht Nitratwerte erhöhen.
7. Warum Nitrat problematisch ist – besonders im Aquarium
Nitrat:
- erhöht Bleaching-Anfälligkeit
- hebt Stickstofflimitierung der Symbionten auf
- führt zu Phosphatlimitierung
- destabilisiert Thylakoidmembranen
- steigert ROS-Produktion
- destabilisiert das Mikrobiom
8. Urea – der ideale Stickstoff für Korallen im Aquarium
8.1 Urea → Urease → NH₃ + CO₂
- NH₃ neutralisiert H⁺ → pH steigt am Kalzifikationsort
- CO₂ liefert anorganischen Kohlenstoff
- CO₂ steigert Photosynthese
8.2 Nickel + Urea
Nickel ist der Cofaktor der Urease.
Nickel + Urea → maximale Kalzifikation & Photosynthese.
Nickel ist als zentraler Cofaktor der Urease unverzichtbar, weil nur bei einer stabilen Nickelkonzentration von etwa 4 µg/L die Urease der Korallen und ihres Mikrobioms maximal aktiv ist und dadurch die Urea‑Spaltung, die Stickstoffversorgung, die CO₂‑Bereitstellung für die Photosynthese sowie die Kalzifikation optimal funktionieren.
9. Urea-Aufnahme bei Algen – wichtig für Aquarianer
Algen können Urea aufnehmen, aber:
- Korallen und Bakterien sind schneller
- moderate Dosierung → Algen profitieren kaum
- erst Überdosierung → Algenwachstum steigt
10. Vergleich der Stickstoffquellen
| Stickstoffquelle | Energiebedarf | Risiko | Wirkung |
|---|---|---|---|
| Nitrat | hoch | ROS, Stress | ineffizient |
| Ammonium | niedrig | gering | bevorzugt |
| Urea | moderat | sehr gering | stabil, wachstumsfördernd |
| Aminosäuren | niedrig | gering | Gewebeaufbau |
11. Ammoniumbicarbonat vs. Ammoniumchlorid
Ammoniumchlorid
- senkt pH
- erzeugt NH₄-Peaks
- keine Pufferwirkung
- erhöht Cyanorisiko
Ammoniumbicarbonat
- stabilisiert pH
- setzt Ammonium gleichmäßiger frei
- liefert Hydrogencarbonat für Kalzifikation (funktional wie „Carbonat“, nur eine Stufe protonierter)
- synergiert perfekt mit Urea
Ammoniumbicarbonat liefert Ammonium + Pufferung + Carbonat.
Ammoniumchlorid liefert nur Ammonium.
12. Stickstoffmanagement heißt: Fluss steuern, nicht Nitratwerte erhöhen
Dies ist der wichtigste Satz der modernen Meeresaquaristik.
Korallen benötigen keinen hohen Vorrat an Stickstoff, sondern einen kontinuierlichen, kontrollierten Fluss in physiologisch sinnvoller Form.
Stickstoff ist im Holobionten kein statischer Wert, sondern ein dynamischer Prozess.
Ein Nitratwert ist lediglich ein Momentfoto, aber kein Abbild des tatsächlichen Stickstoffgeschehens.
Wenn Stickstoff hingegen in der richtigen Form (NH₄⁺ + Urea) und in kleinen Mengen kontinuierlich bereitgestellt wird:
- bleiben Symbiodiniaceae stickstofflimitiert
- erhält die Koralle genug Stickstoff für Wachstum
- bleibt das Mikrobiom balanciert
- wird die Kalzifikation gefördert
- bleibt die pH-Mikrozone stabil
Korallen brauchen Fluss, nicht Vorräte.
Stickstoffmanagement heißt: Den Weg des Stickstoffs steuern – nicht den Nitratwert.
13. Praktische Umsetzung im Aquarium
13.1 Mischung 4:1

Die 4:1‑Mischung bedeutet, dass vier Teile Ammoniumbicarbonat (NH4HCO3) und ein Teil Urea (CH4N2O) kombiniert werden, um Korallen gleichzeitig mit sofort verfügbarem Ammonium und langsam freigesetztem Urea‑Stickstoff in einem stabilen, symbiosefreundlichen Verhältnis zu versorgen.
Die verwendeten Salze sollten grundsätzlich in mindestens Lebensmittel‑ oder idealerweise Ph. Eur.‑Qualität vorliegen, da diese Qualitäten eine Reinheit von meist über 99 % gewährleisten und frei von störenden Schwermetallen sowie organischen Nebenbestandteilen sind.
Beim Ansetzen der Lösung sollte zuerst das Ammoniumbicarbonat vollständig in Osmosewasser gelöst werden, da es nur mäßig löslich ist (ca. 220 g/L bei 20 °C) . Die wässrige Lösung riecht leicht nach Ammoniak – also „ammoniakalisch“ . Erst danach wird der Harnstoff zugegeben, der mit einer Löslichkeit von über 1000 g/L sehr leicht vollständig in Lösung geht.
- 20 g Ammoniumbicarbonat
(auch genannt Ammoniumhydrogencarbonat; M = 79,06 g/mol; CAS Nummer: 1066-33-7) - 5 g Urea
(M = 60,06 g/mol; CAS Nummer: 57-13-6) - in 1 Liter Osmosewasser
Wer anstatt Ammoniumbicarbonat das Ammoniumcarbonat (CH8N2O3 ; M = 96,09 g/mol) verwendet, muss die eingesetzte Menge anpassen: Für die gleiche Ammonium‑Menge entsprechen 20 g Ammoniumbicarbonat etwa 12,1 g Ammoniumcarbonat.
13.2 Dosierung
- 2,5 mL pro 100 Liter täglich; die Dosierung sollte während der Hauptbeleuchtungsphase und in einer einzigen Gabe erfolgen.
Bei dieser Menge entstehen rechnerisch 0,19 mg/L Ammonium, was – falls nicht unmittelbar biologisch verbraucht – theoretisch 0,65 mg/L Nitratäquivalent entsprechen würde. - Bei der Dosierung von 2,5 mL pro 100 Liter entsteht zusätzlich ein minimaler KH‑Anstieg von etwa 0,03 °dKH, der aus der Hydrolyse des Ammoniumcarbonats resultiert.
13.3 Cycling / Start
- 5 mL pro 100 Liter einmalig
14. DON/DOC-Kontrolle
Für die präzise Kontrolle des gelösten organischen Stickstoffs (DON) und des gelösten organischen Kohlenstoffs (DOC) eignet sich ausschließlich die TOC/TNb‑Analyse. Optische Verfahren wie SAC/SAK‑Messungen liefern nur grobe Trends und können die organische Stickstofffraktion nicht zuverlässig erfassen. Die TOC/TNb‑Analyse erlaubt dagegen eine quantitative Bewertung der gesamten organischen Belastung und zeigt, ob der Stickstofffluss stabil bleibt oder sich durch Überdosierung verschiebt.
Die TOC/TNb‑Analyse ist ein kombiniertes Verfahren zur Bestimmung des Gesamtgehalts an organischem Kohlenstoff (TOC) und des Gesamtstickstoffs (TNb) in Wasserproben. Dabei wird die Probe bei hoher Temperatur oxidiert; der entstehende CO₂‑ und NO‑Gehalt wird anschließend gemessen. So lässt sich die Summe aller gelösten organischen Verbindungen quantitativ erfassen – unabhängig davon, ob sie farblos, komplex oder biologisch aktiv sind.
Vorteile:
- Erfasst DON und DOC vollständig, nicht nur optisch aktive Stoffe.
- Unabhängig von Farbe und Trübung der Probe.
- Liefert präzise, reproduzierbare Werte für Belastung und Stoffumsatz im Aquarium.
Damit ist die TOC/TNb‑Analyse die einzige Methode, die den organischen Stickstofffluss im System wirklich quantifizierbar macht – und damit unverzichtbar für eine wissenschaftlich fundierte Kontrolle von DON/DOC.
15. Unterschiede im Stickstoffmanagement bei SPS und LPS
15.1 Grundlegende Unterschiede im Stoffwechsel
- SPS‑Korallen
Hohe Oberflächen‑/Volumen‑Ratio, kleine Polypen, stark lichtgetrieben, hoher Umsatz in Symbiodiniaceae. Sie reagieren sehr sensibel auf Schwankungen im Stickstofffluss, Phosphatlimitierung und ROS‑Stress. SPS benötigen einen sehr gleichmäßigen, fein dosierten Stickstofffluss (Urea + Ammonium), keine Peaks. - LPS‑Korallen
Größere Polypen, stärker heterotroph, können mehr partikuläres Futter und gelöste organische Stoffe direkt nutzen. Sie sind etwas toleranter gegenüber Schwankungen, profitieren aber ebenfalls von DON‑dominierter Versorgung.
15.2 SPS – hohe Symbiontenkontrolle, hohe Fluss‑Sensitivität
Bei SPS ist die Stickstofflimitierung der Symbiodiniaceae besonders kritisch:
- Zu wenig Stickstoff → blasse Farben, Wachstumsstopp, „ausgehungerte“ Kolonien.
- Zu viel Nitrat → Symbiontenüberwuchs, Phosphatlimitierung, Bleaching‑Risiko.
Für SPS gilt daher:
- Urea + Ammonium in kleinen, täglichen Dosen
- keine hohen Nitratwerte als „Sicherheitsreserve“
- Phosphat nicht unter die Nachweisgrenze drücken
- Nickel im Bereich um ca. 4 µg/L halten, damit Urease voll arbeitet.
SPS zeigen sehr schnell, ob der Stickstofffluss passt: Polypenbild, Wachstumsspitzen, Gewebedicke und Farbtiefe sind direkte Indikatoren.
15.3 LPS – mehr Heterotrophie, aber gleiche Prinzipien
LPS können:
- mehr Futterpartikel direkt aufnehmen,
- stärker von Aminosäuren und gelösten organischen Stoffen profitieren,
- Stickstoff teilweise „direkter“ aus dem Wasser holen.
Trotzdem gelten die gleichen Kernprinzipien:
- DON vor DIN (Urea, Aminosäuren vor Nitrat)
- kontinuierlicher Fluss statt Peaks
- keine chronisch hohen Nitratwerte
- Phosphat nicht zu tief ziehen, sonst Stress und Gewebsverlust.
LPS verzeihen Fehler etwas länger, aber wenn sie reagieren, ist der Schaden oft massiver (Gewebsablösungen, Rezession).
15.4 Praktische Konsequenzen für gemischte Becken (SPS + LPS)
In gemischten Riffaquarien mit SPS und LPS ist das Ziel:
- SPS‑tauglichen Stickstofffluss (fein, kontinuierlich, DON‑dominiert)
- LPS‑gerechte Fütterung (partikulär + gelöste Organik)
- moderate Nährstoffwerte:
- Nitrat eher niedrig, aber nicht null
- Phosphat messbar, aber nicht hoch
- Urea + Ammonium 4:1‑Mischung als Basisversorgung
- zusätzliche Fütterung (z. B. feines Frostfutter, Partikel) vor allem für LPS.
16. Fazit für die Meeresaquaristik
Die moderne Forschung zeigt eindeutig:
- Nicht Nitrat erhöhen – Stickstoffform optimieren
- Urea + Ammonium = natürliche, kontrollierte Versorgung
- Nickel als Cofaktor nutzen
- Phosphat nicht zu niedrig halten
- Ziel: stabile, gesunde, wachstumsfördernde Symbiose
Stickstoffmanagement 2.0 ist der Schlüssel zu farbintensiven, widerstandsfähigen und schnell wachsenden Korallen im Aquarium.

 Patrick Gast
Patrick Gast